3.3Les cycles des nutriments dans les systèmes agrisylvicoles
En matière d’efficacité de recyclage des nutriments, on pourrait croire que les systèmes agroforestiers se situent entre les forêts naturelles (cycle des nutriments fermé) et les systèmes agricoles (cycle des nutriments ouvert). C’est partiellement vrai. Les designs des systèmes agrisylvicoles comme les cultures intercalaires sont spécialement pensés de façon à maximiser la rétention des nutriments dans le système, voire à augmenter les sources de nutriments. Dans une culture intercalaire, par exemple, on utilise fréquemment les espèces d’arbres capables de fixer l’azote atmosphérique. Sous un scénario semblable, les cultures intercalaires peuvent accumuler de l’azote et le rendre disponible pour la culture associée. La disponibilité en azote dans ce genre de système peut dépasser celle d’une forêt qui n’abrite pas d’espèces fixatrices de l’azote atmosphérique. De plus, l’espacement entre les rangées d’arbres est choisi afin d’obtenir les meilleures conditions de croissance pour la culture. Pour éviter que les arbres soient en compétition pour l’eau et la lumière, les rangées d’arbres ne doivent pas être trop rapprochées. Toutefois, pour que l’effet fertilisant de la litière comme les feuilles des arbres (un apport annuel de matière organique riche en nutriments et disponible pour les microorganismes décomposeurs) se manifeste sur l’ensemble de la portion de terrain dédiée à l’agriculture, les rangées d’arbres ne doivent pas être très éloignées comme c’est le cas pour les haies brise-vent. Enfin, dans une culture intercalaire, la présence d’arbres favorise aussi les apports primaires en nutriments par filtration de particules atmosphériques (canopée) et par altération des minéraux (exsudats des racines). L’ampleur de ces mécanismes dépend évidemment de l’espèce d’arbre utilisée ainsi que de la densité des arbres dans le paysage, laquelle est déterminée par l’espacement entre les rangées d’arbres et l’espacement des arbres le long des rangées.
Aussi, pour que le système racinaire réduise les pertes de nutriments par lessivage sous la culture agricole, il faut que le système racinaire puisse atteindre les eaux de percolation sous les cultures agricoles. Pour ce faire, on sélectionne des espèces d’arbres avec des systèmes racinaires qui peuvent se développer latéralement sous celui de la culture, du moins en partie. Cela permet le prélèvement des nutriments qui percolent dans le sol et qui sont en excès à la demande de la culture. Dans un système idéal où on évite la compétition interspécifique pour les ressources, l’eau et les nutriments ne sont pas prélevés par les arbres à partir de la même zone que la culture. On cherche plutôt à développer un système par lequel les racines des arbres puisent l’eau et les nutriments en dessous du système racinaire de la culture agricole. En écologie, on parle alors de complémentarité (du concept théorique de la niche écologique). Dans une culture intercalaire bien conçue, les pertes de nutriments par lessivage sont beaucoup plus petites qu’en monoculture. Cela dépend des espèces d’arbres utilisées et de l’espacement choisi.
Quant au recyclage de nutriments, la figure 13 identifie bien les avantages d’une forêt ou d’une culture intercalaire comparativement à une monoculture agricole.

Figure 13. Schématisation du cycle des nutriments en forêt, en agriculture et en agroforesterie.
Source : Adapté de Nair (1984).
Les cultures intercalaires et beaucoup d’autres systèmes agrisylvicoles cherchent à promouvoir le recyclage des nutriments par l’un ou plusieurs des mécanismes suivants : (1) la fixation de l’azote atmosphérique, (2) l’apport de matière organique (litière) riche en nutriments pouvant être mobilisés par les microorganismes (ex. feuilles), (3) le prélèvement accru de nutriments par les racines des arbres sous le système racinaire de la culture agricole (réduction des pertes par lessivage), (4) l’augmentation des flux primaires de nutriments comme l’altération des minéraux et la filtration des particules atmosphériques et (5) une meilleure protection des sols contre l’érosion. Pour la suite de cette section, nous mettrons l’accent sur les quatre premiers mécanismes. Nous n’aborderons pas davantage le point 5, car nous en discuterons en détail dans le module traitant des systèmes environnementaux.
3.3.1La fixation de l’azote atmosphérique par les arbres
L’usage des espèces fixatrices de l’azote atmosphérique est très commun en agriculture ainsi qu’en agroforesterie. Puisque l’azote joue un rôle très important dans le métabolisme des plantes (il est le constituant principal des protéines et conséquemment de la matière vivante), plusieurs techniques sont utilisées aujourd’hui pour contrer l’appauvrissement des sols en azote. En effet, les fertilisants à base d’azote ou encore les espèces de plantes capables de fixer biologiquement l’azote atmosphérique (en jachère ou en culture intercalaire) sont couramment utilisés pour enrichir les sols et, ainsi, maximiser les rendements des récoltes à venir.
La majorité des plantes ne peuvent pas prélever l’azote sous sa forme gazeuse (N2). L’azote doit être assimilé sous ses formes minérales, soit l’ion ammonium (NH4+) et l’ion nitrate(NO3-). Pour en arriver à NO3-, l’azote contenu dans la matière organique est minéralisé par les microorganismes à travers une série de processus d’oxydation qui se traduit par la perte d’électrons de l’atome d’azote. C’est ce qu’on appelle la nitrification. Les ions NO3 sont ensuite facilement assimilés par les plantes en échange d’ions hydroxyle (OH-). La nutrition azotée des plantes est assurée surtout par NO3; toutefois, les plantes qui exploitent les milieux acides ont développé la capacité de prélever NH4 en échange de proton (H+), étant donné la faible activité des bactéries nitrifiantes sous de telles conditions. La nitrification est donc très limitée et le NH4 domine la solution de sol.
Toutefois, les légumineuses (ou familles des fabacées) et d’autres espèces de plantes (dites actinorhiziennes) ont l’avantage de ne pas nécessiter un accès direct aux ions NH4+ et NO3– pour satisfaire leurs besoins en azote. Pour ces espèces, des bactéries fixatrices d’azote développent des associations symbiotiques avec le système racinaire des plantes. Les rhizobiums sont les bactéries fixatrices d’azote qui développent des symbioses avec les légumineuses sous la forme de nodosités (figure 14), alors que les actinobactéries (ou bactéries actinomycètes) du genre Frankia développent des associations avec des arbres et des arbustes comme l’aulne, l’argousier et les éricacées (figure 15). Les nodosités sont appelées des actinorhizes.

Figure 14. Nodosités (Rhizobium) sur les racines d’un plant de soja, une légumineuse.
Source : https://mytakeontoday.wordpress.com/tag/soybean-root-nodule/

Figure 15. Actinorhizes (Frankia) sur les racines d’un aulne rouge (Alnus rubra).
Source : https://hsc.forestry.oregonstate.edu/red-alder-physical-characteristics
C’est à partir d’un poil racinaire que les bactéries développent leurs associations avec la plante. Le poil est, en premier lieu, infecté par les bactéries à l’aide de signaux que la plante achemine aux bactéries pour signaler que le passage est ouvert (figure 16). Avec d’autres signaux envoyés par les bactéries à destination, le poil s’enroule pour enrober et fixer les bactéries. C’est à partir de cette boucle que se forme un cordon d’infection jusqu’à la couche superficielle (cortex) de la racine (figure 16). La nodosité se forme à partir du cordon à la suite de la division cellulaire. C’est alors que les bactéries, maintenant multipliées, s’insèrent dans la nodosité. Une fois suffisantes, les bactéries arrêtent de se multiplier pour ne pas faire gonfler davantage la nodosité. Le coût énergétique pour que la plante bâtisse les nodosités est très grand.
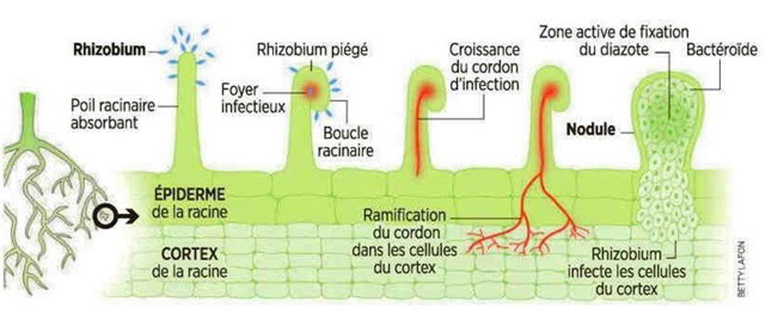
Figure 16. La formation d’un nodule à partir d’une infection par les rhizobiums.
Source : Adapté de https://planete.gaia.free.fr/animal/symbiose/bacteplantes.html
Par des actions biochimiques complexes, ces usines bactériennes (c’est-à-dire les nodosités) produisent la nitrogénase, une enzyme qui favorise la synthèse de l’ammoniac (NH3) à partir de l’azote gazeux dans le sol (N2). La synthèse de NH3 se fait par un processus de réduction se traduisant par le gain d’électrons de l’atome d’azote. Par la suite, le NH3 est transformé en formes utilisables pour les plantes (ex. acides aminés).
Bien entendu, les taux de fixation de l’azote atmosphérique varient d’une espèce à l’autre. Ils varient même selon la provenance d’une même espèce. Par exemple, une population d’aulnes rouges établie sur un sol intrinsèquement pauvre en azote aura vraisemblablement des taux de fixation plus élevés qu’une population évoluant sur des sols riches en azote, parce que l’investissement en énergie pour former les nodosités est une mauvaise stratégie pour les aulnes sur sols riches. La fixation en azote par certaines forêts varie entre 50 et 300 kg N ha-1 an-1, voire 500 kg N ha-1 an-1 dans certains cas extrêmes. En comparaison, les précipitations fournissent entre 10 et 15 kg N ha--1 an-1 au sud du Québec (figure 17). C’est plus élevé dans les régions très polluées (jusqu’à 40 kg N ha-1 an-1). Si l’on considère que l’atmosphère est la seule source nette d’azote pour les écosystèmes terrestres et que les précipitations représentent généralement une petite source en régions éloignées de la pollution, la fixation de l’azote atmosphérique devient une source importante pour plusieurs écosystèmes forestiers.
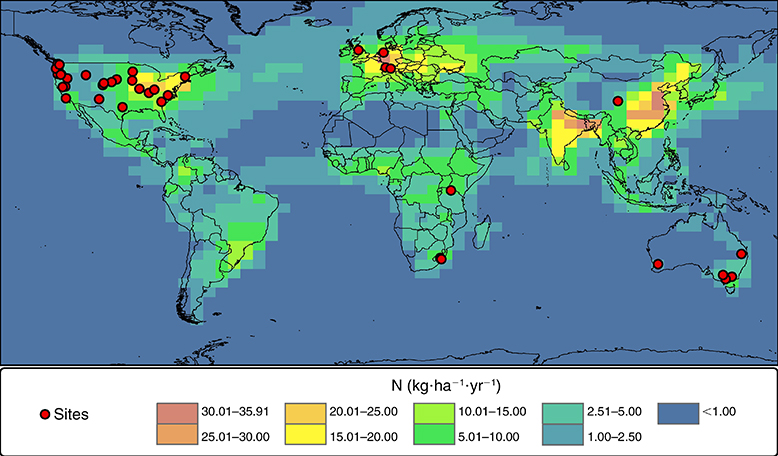
Figure 17. Taux de déposition atmosphérique d’azote sur la surface terrestre.
3.3.2L’apport de matière organique riche en nutriments par la litière foliée
Les légumineuses prennent toutes sortes de formes. Elles sont des arbres, des arbustes, des lianes et des herbacées. Certaines espèces sont rustiques aux zones froides, alors que d’autres sont rustiques aux zones tropicales. Les légumineuses comptent plusieurs espèces d’intérêt économique comme le haricot, le pois, la lentille, le pois chiche, le soja, la fève, l’arachide, la luzerne, la réglisse et le trèfle. Par leurs fortes teneurs en protéines et en acides aminés, elles jouent un rôle important dans l’alimentation humaine.
Au cours des années, certaines espèces d’arbres et d’arbustes sont devenues des incontournables dans les paysages agroforestiers parce qu’elles ont la capacité d’enrichir directement le sol en azote en fixant biologiquement le N2 atmosphérique. On s’en sert souvent pour les cultures intercalaires, les jachères rotationnelles forestières, les cultures pérennes ombragées et comme arbres aléatoires sur les terrains agricoles. Toutefois, leur capacité à enrichir le sol ne dépend pas uniquement des taux de fixation de l’azote atmosphérique. Elle dépend aussi de la qualité de leurs litières, notamment les feuilles. On peut dire qu’une feuille qui se décompose rapidement lorsqu’elle atteint le sol relâchera plus rapidement les nutriments qu’elle contient. Si cette feuille est riche en azote, elle relâchera une bonne quantité d’azote que la récolte agricole pourra utiliser pour sa croissance.
Les taux de décomposition des feuilles dépendent de deux facteurs principaux, soit le rapport carbone/azote (C/N) et l’acidité du sol. Dans l’ensemble, plus le rapport C/N d’une feuille est faible, plus elle se décompose rapidement. Cela s’applique aux différentes matières organiques. Le carbone est utilisé par les microorganismes à des fins énergétiques (2/3) et pour maintenir leur constitution (1/3). L’azote, quant à lui, n’est utilisé par les microorganismes que pour maintenir leur constitution. Puisque l’azote est l’élément le moins abondant et le plus limitant pour les microorganismes du sol, on utilise le rapport C/N plutôt que le rapport N/C. Pour une feuille, par exemple, un rapport C/N typique de 20 serait exprimé par un rapport N/C de 0,05.
Une bactérie a un rapport C/N moyen de 6 à 10. Si le rapport C/N de la feuille est de 10 à 15, l’activité des microorganismes sera à son maximum, car la feuille répond très bien à leurs exigences en azote. En conséquence, la vitesse de décomposition de la feuille est très rapide. On dit alors que c’est une matière organique de qualité, car elle mobilise rapidement les nutriments qu’elle contient lorsqu’elle parvient à la surface du sol. Toutefois, les espèces d’arbres et d’arbustes qui produisent des feuilles avec de tels rapports C/N sont assez rares. Les feuilles des arbres et arbustes ont typiquement des rapports C/N variant entre 15 et 35. Avec un rapport C/N de 15 à 25, les besoins en azote ne sont pas parfaitement comblés. L’activité des microorganismes et les taux de décomposition de la feuille ralentissent. Un rapport C/N de 25 à 35 signifie que l’azote est limitant (25) à très limitant (35). L’activité des microorganismes est alors faible à très faible. Les forêts de conifères génèrent des litières d’aiguilles qui sont acidifiantes et caractérisées par des rapports C/N de plus de 30. Sous les conditions de sol acides (pH de 4,5 ou moins) et une mauvaise qualité de litière, l’activité microbienne (spécialement les bactéries) et les taux de décomposition sont au ralenti. En fait, les champignons plutôt que les bactéries dominent la flore microbienne sous de telles conditions. Toutefois, les champignons n’ont pas la même efficience à décomposer la matière organique que les bactéries. De ce fait, d’épaisses couvertures mortes (humus) se forment à la surface du sol. Ce type de couverture morte est appelé « mor » (figure 18).

Figure 18. Épaisse couverture morte (mor) de la forêt boréale en Suède.
Au contraire, quand la litière des arbres est de bonne qualité (rapport C/N de 15 ou moins) et que le sol n’est pas trop acide (pH de 5,5 ou plus), on peut s’attendre à une activité bactérienne et des taux de décomposition de la matière organique élevés. S’il y a présence d’une microfaune fouisseuse (ex. vers de terre), la matière organique est incorporée au sol minéral et elle est décomposée in situ par les bactéries pour former un « mull » (figure 19). Ce type de sol, caractérisé par des complexes (ou agrégats) organominéraux stables, est très convoité pour l’agriculture en raison de sa résistance à l’érosion et de sa capacité à retenir l’eau et les nutriments.

Figure 19. Exemple de mull exploité pour l’agriculture.
Source : https://www.digopaul.com/fr/english-word/chernozem.html
L’acidité du sol et le rapport C/N des feuilles sont de bons indicateurs servant à prédire les taux de décomposition de la litière. Toutefois, les feuilles de certaines espèces se décomposent lentement malgré les rapports C/N assez bas et la présence de bactéries. Les scientifiques ont récemment découvert que la teneur en lignine contrôle aussi les taux de décomposition des feuilles. La lignine est l’une des principales composantes du bois, variant de 23 % (feuillus) à 27 % (conifères). La cellulose (41 à 43 %), l’hémicellulose (20 à 30 %) et l’eau sont les trois autres composantes du bois. La lignine est une biomolécule qui résiste à la décomposition par les microorganismes. Elle est aussi présente dans les feuilles, variant de 3 à 5 %. Par exemple, les feuilles de plusieurs espèces d’acacias sont riches en lignine. Malgré un rapport C/N des feuilles d’environ 18 chez l’Acacia augustissima, elles sont plutôt récalcitrantes à la décomposition, compte tenu de leur forte teneur en lignine. Le tableau 1 présente le lien entre la capacité d’une espèce à fixer l’azote atmosphérique, le rapport C/N, la teneur en lignine et le taux de décomposition des feuilles d’espèces arborées et arbustives couramment utilisées en agroforesterie.
Tableau 1. Qualité et vitesse de décomposition de la litière d’espèces végétales utilisées en agroforesterie, certaines capables de fixer l’azote atmosphérique, d’autres incapables.
| Espèces | Fixation de l’azote | Rapport C/N | Teneur en lignine | Taux de décomposition |
|---|---|---|---|---|
| Gliricia sepium | Oui | 10 à 20 | + | Rapide |
| Acacia augustissima | Oui | 18 | +++ | Lent |
| Sesbania sesban | Oui | 15 à 16 | + | Rapide |
| Grevilia robusta | Non | 32 | +++ | Lent |
| Dactyladenia barteri | Non | 25 à 30 | ++++++ | Lent |
| Senna reticulata | Non | 18 | + | Moyen |
| Myroxylon balsamum | Oui | 16 | + | Moyen |
Source : Adapté de Nair (1998).
Enfin, la température et la teneur en eau du sol sont deux autres facteurs qui influencent la décomposition de la feuille. L’activité des microorganismes augmente avec la hausse de la température et de la teneur en eau du sol. Toutefois, des températures trop élevées, normalement accompagnées par des stress hydriques, finissent par nuire à l’activité microbienne. De façon similaire, la saturation et la stagnation de l’eau dans le sol diminuent les niveaux d’oxygène, ce qui nuit considérablement à l’activité des microorganismes.
En agroforesterie, il faut donc considérer un éventail de facteurs pour mieux prédire la vitesse de décomposition d’une feuille et, de ce fait, le taux de mobilisation des nutriments. Il faut aussi considérer le flux de litière à la surface du sol que peuvent apporter les arbres et les arbustes d’un système agrisylvicole pour mieux évaluer l’impact de la litière sur la qualité du sol, notamment sa teneur en carbone.
On peut modéliser l’effet de la litière foliée assez simplement en utilisant une constante de décomposition (k), laquelle tient compte théoriquement de tous les facteurs discutés précédemment, et un flux de litière (FL). Si on présume des valeurs de k de 0,1, 0,2 et 0,3 et un flux de 1000 kg ha-1 an-1, on obtient trois teneurs en carbone organique du sol (COS) de la façon suivante :
1re année de COS = FL – (k × FL)
2e année de COS = [1re année de COS + FL] – [k × (1re année de COS + FL)]
et ainsi de suite… Une valeur k de 0,1 signifie que seulement 10 % de la matière organique est décomposé annuellement, alors qu’une valeur de 0,3 signifie que 30 % sont décomposés. À l’état d’équilibre (plafonnement de la courbe), on peut voir dans la figure 20 que le COS est plus faible (2400 kg ha-1) si la décomposition de la litière est plus rapide (k = 0,3) et que le COS est plus élevé (8700 kg ha-1) si la décomposition de la litière est plus lente (k = 0,1). Cela signifie que le sol accumule plus de COS et libère moins de nutriments contenus dans la litière foliée si la décomposition est plus lente, alors que le sol accumule moins de COS et libère plus de nutriments contenus dans la litière si la décomposition est rapide. Il faut aussi plus de temps pour atteindre l’état d’équilibre de 8700 kg COS ha-1 avec un k de 0,1 que pour atteindre 2400 kg COS ha-1 avec un k de 0,3 (figure 20).
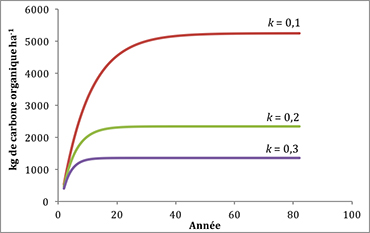
Figure 20. Évolution du carbone organique du sol selon un flux de litière de 1000 kg ha-1 an-1 et des constantes de décomposition de 0,1, 0,2 et 0,3. Cliquez ici pour consulter le fichier Excel.
Si le rendement des arbres diffère d’un site à l’autre parce qu’il y a un gradient de disponibilité en eau, par exemple, le flux de litière peut passer de 1000 kg ha-1 an-1 (site sec et improductif) à 3000 kg ha--1 an-1 (site arrosé et productif). Si on présume un k de 0,1, le temps nécessaire pour atteindre l’état d’équilibre de COS est le même, peu importe le flux de litière choisi (figure 21). Toutefois, la valeur maximale de COS atteint 27 000 kg ha-1 sous un flux de 3000 kg ha-1, alors qu’elle n’est que de 9000 kg ha-1 sous un flux de litière de 1000 kg ha-1 (figure 21).
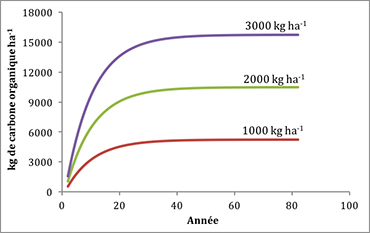
Figure 21. Évolution du carbone organique du sol selon une constante de décomposition de 0,1 et des flux de litière de 1000, 2000 et 3000 kg ha-1 an-1. Cliquez ici pour consulter le fichier Excel.
Une lente décomposition de la litière a le mérite d’augmenter le COS. Il s’agit là d’un bénéfice direct pour le sol, car de fortes teneurs en carbone favorisent la formation d’agrégats stables avec les particules argileuses. Ces agrégats améliorent la structure du sol et sa résistance à l’érosion. Un sol plus riche en carbone aura aussi une capacité de rétention en eau et un contenu en nutriments plus élevés qu’un sol pauvre en carbone. L’augmentation des teneurs en COS est ainsi perçue positivement en agriculture.
La plus grande critique d’un système agrisylvicole qui incorpore des espèces d’arbres et d’arbustes caractérisées par des litières foliées récalcitrantes pourrait venir du producteur agricole qui cherche à fertiliser immédiatement ses cultures par le flux annuel de litière. En effet, la lente décomposition permet d’accumuler le carbone dans le sol. Par le fait même, les autres nutriments dans la litière s’accumulent aussi dans le sol. Ces nutriments ne sont toutefois pas nécessairement dans des formes facilement assimilables. Ceux-ci doivent être minéralisés davantage par les microorganismes avant d’être mobilisés sous des formes disponibles pour la culture. Au contraire, une litière facilement décomposable ne favorisera pas l’accumulation de carbone et de nutriments dans le sol. Le carbone sera utilisé par les microorganismes pour de l’énergie et ensuite perdu en CO2 vers l’atmosphère ou par lessivage sous forme de carbone organique dissous, alors que les nutriments seront vite minéralisés et rendus directement disponibles pour la culture.
Ainsi, le producteur agricole peut introduire des espèces d’arbres et d’arbustes avec des litières de grande qualité et d’autres de moins bonne qualité, de façon à fertiliser sa culture (k élevé) et à bâtir des réserves de carbone et d’autres nutriments dans le sol (k plus faible), respectivement. D’après la figure 22, il est possible de faire correspondre la mobilisation des nutriments à la demande en nutriments de la culture. En (a), une litière foliée de grande qualité mobilisera les nutriments à des taux plus élevés que la demande de la culture. Il y aura donc de larges pertes en nutriments par lessivage. En (b), une litière de faible qualité mobilisera les nutriments à des taux plus faibles que la demande de la culture. La culture expérimentera alors des déficiences nutritionnelles. En (c), le mélange d’espèces fournissant autant des litières de bonnes que de faibles qualités permet d’éliminer complètement les déficiences nutritionnelles et de limiter les pertes de nutriments par lessivage. Ce genre de compromis est souvent visé en agroforesterie. En (d), on simule l’absence de litière, ce qui se traduit par de faibles pertes par lessivage en début de culture, mais des déficiences plus tard lorsque les besoins nutritionnels de la culture augmentent.
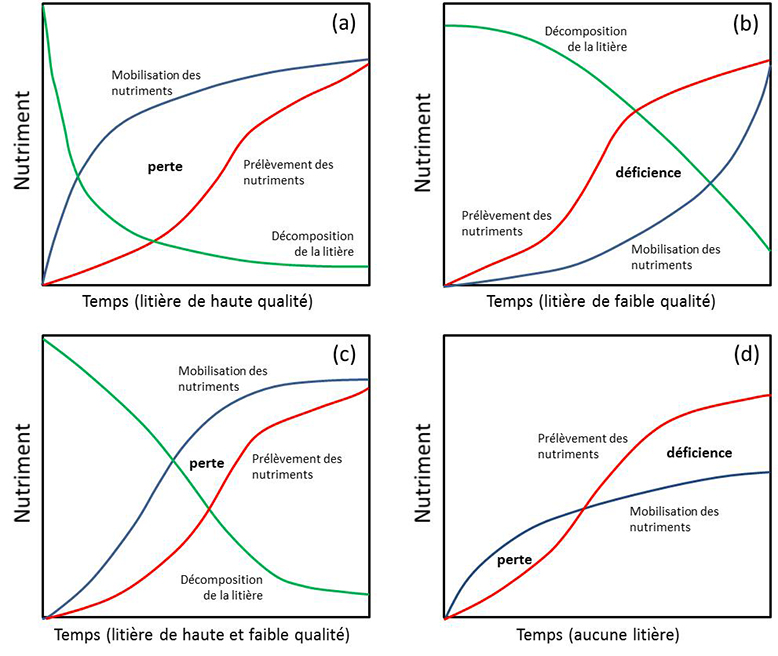
Figure 22. Perte et déficience de nutriments selon la qualité de la litière. Voir les explications dans le texte ci-dessus.
Source : Adapté de Myers et al. (1994).
3.3.3 Les arbres : une source toute spéciale de nutriments dans les écosystèmes
Les résultats d’une recherche menée par le professeur responsable de ce cours, Nicolas Bélanger, vous aideront à mieux visualiser comment les arbres peuvent parvenir à augmenter la disponibilité des nutriments dans le sol en réduisant les pertes par lessivage et en augmentant les flux primaires de nutriments comme l’altération des minéraux et la filtration des particules atmosphériques (Bélanger et al., 2004). Cette recherche a été réalisée dans une plantation d’une trentaine d’années d’épinettes de Norvège (Picea abies [L.] Karst.), à Valcartier, à une vingtaine de kilomètres au nord de la ville de Québec. Dans cette plantation, on a testé la croissance d’épinettes de Norvège de différentes provenances, c’est-à-dire des épinettes de la même espèce, mais que l’on retrouve dans différentes régions (ex. une épinette du nord de la Norvège, une du sud de la Suède, une de la Russie, une du nord de l’Allemagne, etc.). En testant les provenances, on a cherché à identifier celles qui performent le mieux sous le climat québécois. Dans cette plantation, selon les provenances, il y a des épinettes de Norvège qui poussent très bien et d’autres qui poussent beaucoup moins bien.
En théorie, puisqu’il s’agit de la même espèce, seules les différences de croissance entre les provenances productives et les provenances les moins productives devraient avoir un impact sur la disponibilité des nutriments du sol. Les autres facteurs influençant le cycle des nutriments (ex. la litière) devraient être les mêmes puisqu’il s’agit de la même espèce. Plus l’épinette de Norvège est productive, plus le prélèvement de nutriments devrait se faire à des taux élevés. Dans ce sens, les sols qui supportent les provenances les plus productives devraient contenir moins de nutriments que les sols qui supportent les provenances les moins productives. C’est exactement cette hypothèse qui a été testée. À la grande surprise de l’équipe de chercheurs, c’est plutôt le contraire qui a été observé (figure 23). Par exemple, les provenances les plus productives prélèvent beaucoup plus de calcium (environ 100 g m-2 dans les aiguilles, les branches et les tiges) que les provenances les moins productives (environ 35 g m-2). Pourtant, les sols des provenances les plus productives contiennent davantage de calcium (environ 130 g m-2) que les sols des provenances les moins productives (environ 100 g m-2).
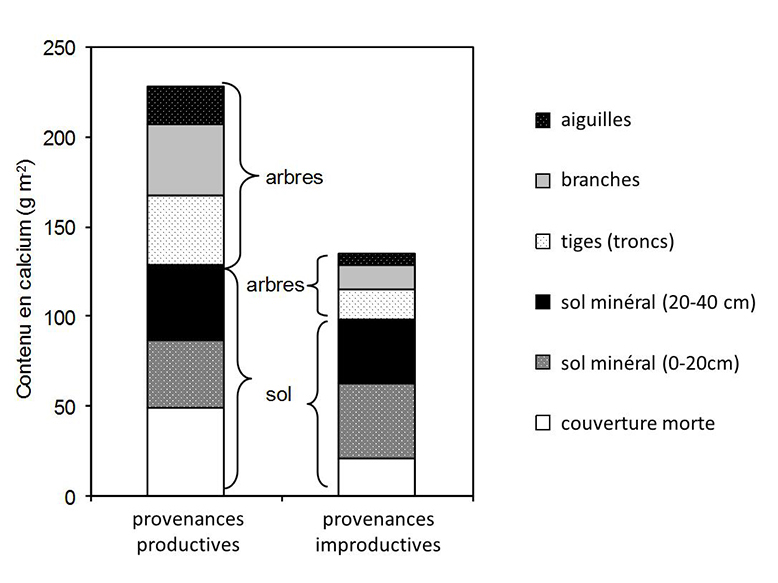
Figure 23. Contenus en calcium du sol sous des provenances d’épinettes de Norvège à productivité contrastée. Les contenus en calcium dans la biomasse sont aussi indiqués.
Source : Bélanger et al. (2004).
Pour que les sols sous les provenances les plus productives contiennent le plus de calcium, c’est que leur croissance accrue génère des sources additionnelles de calcium qui dépassent largement les taux de prélèvement (puits). On peut penser à trois types de flux qui sont augmentés sous une croissance accrue des épinettes. Notamment, les provenances plus productives ont des canopées mieux développées, plus hautes et plus garnies. Ainsi, la canopée des provenances les plus productives filtre plus efficacement les particules atmosphériques, apportant par le fait même du calcium et d’autres nutriments. De façon similaire, les provenances les plus productives ont développé des systèmes racinaires plus complexes pour bien ancrer les arbres et soutenir leurs besoins en eau et en nutriments. Par un système racinaire mieux développé, ces provenances génèrent plus d’exsudats acides, lesquels augmentent les taux d’altération des minéraux du sol et, par conséquent, les taux de mobilisation du calcium. Avec un système racinaire plus imposant et un prélèvement d’eau accru, les provenances plus productives limitent aussi les pertes de nutriments par lessivage. C’est ainsi qu’on retrouve des contenus en calcium plus élevés sous les provenances productives, malgré un prélèvement plus important. L’effet net d’une croissance accrue est l’augmentation des contenus en calcium.